Новый взгляд на патогенез и лечение шизофрении
А.И. Воронов
ФГБНУ «Научно-исследовательский Институт Фундаментальной и Клинической иммунологии» (НИИФКИ), г. Новосибирск
Е-mail: voronov88888@yandex.ru
В основе патогенеза всех типов шизофрении по мнению автора лежат специфические нарушения сна. Дефицит дельта сна порождает большинство клинических симптомов, на которых всегда строился диагноз шизофрении. Псевдогаллюцинации, падение энергетического потенциала, нарушения внимания и мышления, бредовые идеи – не что иное, как вторичные проявления дефицита дельта сна! Психотропные препараты действуют на нейроны в области синапса и, по сути, являются корректорами поведения. Без надежды на исцеление их приходится принимать всю жизнь. Действие нейролептиков практически не влияет на структуру дельта сна, не останавливает процессы нейровоспаления и нейродегенерации. Дефектную симптоматику самые современные нейролептики тоже не отменяют. Представленная модель патогенеза во многом гипотетична и будет совершенствоваться по мере накопления знаний о мозге, но она дает целостную картину развития заболевания и объясняет клиническую эффективность применения цитокинов.
Ключевые слова: патогенез шизофрении, дельта сон, цитокины, лечение
Знание некоторых принципов избавляет от знания многих фактов…
Сто пятьдесят лет назад Дмитрий Иванович Менделеев открыл периодический закон, положив в основу принцип возрастания атомного веса. Это позволило расположить в строгом порядке более 60 известных на тот момент химических элементов. В таблице оставались «пустые» клетки и Менделеев заранее предсказал свойства некоторых, тогда ещё не открытых, веществ. Теперь они заняли «свои» места, подтвердив правильность выбранного принципа.
За 100 с лишним лет изучения шизофрении накопилось множество клинических наблюдений, собранных теперь в МКБ 10 в виде симптомов и синдромов, ни один из которых «не считается патогномоничным» [15]. Их непредсказуемые сочетание и поразительное разнообразие у «лично больных», [1] иногда ставит в тупик даже опытных психиатров, особенно, когда необходимо присвоить диагностический шифр конкретному пациенту. Случается, что обязательных признаков «не хватает», а второстепенных более чем достаточно. Приходится долго наблюдать лично больных в условиях стационара, прежде чем решиться на серьезный диагноз.
Мы хотим предложить вниманию читателей последовательный ряд патогенетических механизмов, способный объединить все это многообразие несколькими принципами. Любопытно, что некоторые из них уже давно лежат на поверхности.
До сих пор основой диагностики шизофрении остается клиническое интервью. Одинакового понимания психиатрами принципов формирования диагноза, выбора лечения и внятного прогноза – по сей день нет. Наиболее красноречиво об этом говорит эксперимент Девида Розенхана проведенный в 1973 г. Восемь психически здоровых граждан (врачей, юристов, психологов) обратились в разные психиатрические клиники с жалобами на голос в голове произносящий слова: «Плюх. Падение. Пустота». В остальном все вели себя спокойно, на вопросы отвечали правильно, тем не менее, все были госпитализированы и «пролечены». Из стационара их не выпускали до тех пор, пока они не «признали себя шизофрениками». Юрист вообще провел там 52 дня. Публикация результатов эксперимента вызвала скандал и поток критики. Большинство психиатров утверждали, что никогда бы не перепутали больных и здоровых людей. Розенхану предложили продолжить эксперимент, он согласился. По больницам прокатилась волна обнаружений пациентов, «засланных» Розенханом. Почти четверть вновь поступивших больных была сочтена симулянтами. Но как оказалось, во второй раз Розенхан никого не посылал, и ему снова удалось всех обмануть.
С тех пор мало что изменилось…
Существует несколько основных подходов в объяснении механизма развития шизофрении.
Пока, среди практикующих психиатров преобладает «дофаминовая» теория, считающая основной причиной шизофрении дисбаланс различных медиаторов: глутамата, серотонина и дофамина. Все терапевтические усилия современной психиатрии направлены на устранение этого дисбаланса, не смотря на то, что доказательств гиперфункции дофаминергических нейронов по сей день – нет! Изначальная догадка о переизбытке дофамина в синапсах нейронов среднего мозга не подтвердилась. Возник вопрос: «Почему, и каким образом помогают нейролептики?» Ответ дал Арвид Карлссон. Он предположил, что для шизофрении характерно нарушение регуляции активности дофаминергических нейронов, а не их гиперфункция. За исследования роли дофамина в мозговой деятельности в 2000 году он получил нобелевскую премию.
Второй подход делает акцент на повышение активности нейронов гиппокампа, фронтальных и височных долей мозга. Такая активность при шизофрении реально регистрируется, хотя ее причины пока не понятны…
Наконец третий подход указывает на структурные изменения нейронов и глии, распад нервных связей и нейродегенерацию. В капитальном исследовании под редакцией Тиганова сказано: «Особенностью шизофрении является снижение реактивности нейроглии: отсутствие пролиферативной реакции астроглии, атрофичность микроглии и регресс олигодендроглии... В очагах выпадения нервных клеток отсутствует заместительный глиоз, свойственный другим патологическим процессам в нервной системе». Более того, оказалось, что функциональный статус микроглиальных клеток тесно связан с состоянием периферической иммунной системы.
Активация микроглии часто существенно опережает нейродегенерацию во времени и может наблюдаться даже в ее отсутствие. Это дает основание предположить, что реактивность микроглии может быть очень важным фактором в патогенезе шизофрении, поскольку имеется большое число работ, в которых были описаны нарушения иммунного статуса при этом заболевании.
Во время острого психотического эпизода меняются пропорции CD4+ и CD8+ Т-лимфоцитов, растет пропорция активированных лимфоцитов, а также аккумуляция макрофагов.
На посмертном материале 40 пациентов, с подтвержденным диагнозом шизофрения, Н.С. Коломеец показала, что больше всего страдают олигодендроциты, особенно в случае преобладания негативной симптоматики. А ведь кроме олигодендроцитов миелин в мозге никакие другие клетки не производят. Следствием этого является демиелинизация аксонов – так называемый симптом «потертых проводов», ведущий к рассеиванию аксонных импульсов. С каждым следующим приступом прогрессивно нарушается ультраструктура астроцитов. Численная плотность и объемная фракция митохондрий в астроцитах достоверно падает по мере увеличения длительности болезни. Практически исчезает рамифицированная и круглая микроглия.
Все три подхода и ряд других теорий пытаются объяснить отдельные механизмы формирования клинических синдромов, но внятно связать все многообразие накопленных фактов с их помощью пока не получается.
Нейрофизиология в последние годы развивается особенно бурно: появились технические возможности непосредственного, визуального наблюдения за работой живого мозга. Например, таким способом была открыта глимфатическая система. Целый ряд открытий произошел в области иммунологии и медицины сна.
Особенно интересно открытие ранее неизвестных функций астроцитов. Оказалось, что астроциты «забирают» поврежденные митохондрии у нейронов и отдают им полноценные – собственные. Как бы «обмениваются энергетическими станциями». Высвобождение астроцитами митохондриальных телец в межклеточное пространство опосредованно кальций-зависимыми механизмами. Кроме того, психиатрам надо учитывать еще один важный момент: астроциты, попавшие в неблагоприятные условия (атакованные антителами), не только перестают снабжать нейроны своими «энергетическими станциями», но и перестают поставлять им лактат молочной кислоты. Обратите внимание это очень важно! В процессе гликолиза, астроциты образуют две молекулы АТФ (аденозинтрифосфорная кислота – универсальный источник энергии для всех биохимических процессов) и лактат молочной кислоты, который, как и митохондрии, перемещают во внеклеточное пространство. Лактат и митохондрии, захватываются нейронами. В нейронах лактат восстанавливается до пирувата и через цикл трикарбоновых кислот на митохондриальной цепи образует 38 молекул АТФ. Для сравнения – сами астроциты, в ходе гликолиза, получают для своих энергетических нужд только две молекулы АТФ. По факту, именно астроциты полностью обеспечивают высокие потребности нейронов в энергии. Если у астроцитов возникают «проблемы» – это немедленно сказывается на нейронах. Астроциты запасают энергию, которую отдают нейронам, проявляя удивительную энергетическую щедрость. После аутоиммунного поражения астроцитов – энергетика мозга ощутимо страдает. Недаром пациенты шизоидного спектра ощущают отсутствие энергии, апатию, быстро утомляются, уходят в себя, погружаются в аутизм. Им не хватает энергии для полноценных социальных контактов.
Интересно, что астроциты при определенных условиях способны превращаться и превращаются в нейроны. Кроме того, астроциты оказались обязательными участниками передачи нейронного импульса. Классическая модель синапса уходит в прошлое. Астроциты окружают пресинаптическую и постсинаптическую мембраны нейрона и на равных участвуют в передаче сигнала. Они выделяют АТФ, ГАМК, серин и другие нейромедиаторы. Модель функционального трехстороннего синапса: аксон – дендрит – астроцит – доказан. Остается доказать, что именно астроциты, в определенных условиях, повышают чувствительность рецепторов к дофамину, особенно в зоне гиппокампа, варолиева моста, обонятельного мозга.
Между собой астроциты «общаются» посредством щелевидных кальциевых каналов, образуя домены. Связь между доменами осуществляется посредством особого типа астроцитов с очень длинными отростками. Такие отростки обнаруженных на астроцитах человека и высших млекопитающих. Ритмическая активность мозга генерируется скоординированным действием нейрональных и глиальных сетей.
«Конечные ножки» асторцитов плотно, без промежутков, окружают все кровеносные сосуды головного и спинного мозга, включая капилляры и анастомозы. В подошвенной мембране «ножек» находятся водные каналы, направленные строго внутрь астроцита. Они представлены белком аквапарин-4. По плотности водных каналов на мембранах «ножек», астроциты приближаются к почкам – органам, интенсивно перекачивающим воду. Все вместе, «конечные ножки» астроцитов составляют мозговую границу гематоэнцефалического барьера. Сосудистую границу составляет плотный эпителий всех мозговых сосудов. Между этими образованиями имеется периваскулярное пространство, по которому движется ликвор. В эндотелии сосудов аквапориновые каналы отсутствуют. Это означает, что жидкость из сосудов не может проникнуть в ткани мозга. Она течет по периваскулярным пространствам и попадает в ткани мозга, только проходя через астроциты.
Астроцитарные «ножки» последовательно, доменами, увеличиваются в объеме, образуя подобие муфты, затем «съеживаются», продвигая ликвор по периваскулярному пространству. В момент «съеживания» ножек просвет между эпителием сосуда и «ножками» астроцитов увеличивается на 60%. Чем не двигатель, недавно открытой, глимфатической системы. Интересно, что участие в управлении этим двигателем во многом принадлежит норадреналину: в состоянии бодрствования содержание норадреналина достоверно повышается, а во время сна снижается, что и приводит к усилению движения жидкости.
Величайший русский исследователь и психиатр, П.Е. Снесарев, еще в пятидесятых годах прошлого века, намного опередил западных коллег в своем утверждении о том, что «в основе патогенеза шизофрении лежит интоксикация специфическими аллергенами эндогенного происхождения». Достижения современной иммунологии показали сходство шизофрении с хроническими аутоиммунными заболеваниями по целому ряду признаков.
С одной стороны налицо активация гуморального звена иммунной системы всего организма, в виде увеличения содержания иммуноглобулинов. С другой – изменение клеточного иммунитета. В течение всего заболевания В-клеточный иммунитет повышен, а в периоды инициации или обострения нарушен баланс цитокинов с преобладанием Тh2.
По данным большинства авторов снижено количество Т-хелперов (Th-1), что ведет к дисбалансу Th-1 / Th-2 системы.
В сыворотке крови больных шизофренией присутствуют специфические антимозговые антитела. Как следствие аутоиммунного патологического процесса, при шизофрении повышена проницаемость гематоэнцефалического барьера. Тем не менее, шизофрению нельзя отнести к классическим аутоиммунным заболеваниям. Мы полагаем, что аутоиммунная атака на астроциты является только пусковым моментом (капсулем, запалом). Дальше патологический процесс течет по иным законам. Да и сам «пусковой момент» может не случиться в течение всей жизни, а сохранится в геноме и передаться следующему поколению. Выброс антител к собственному мозгу и прорыв гематоэнцефалического барьера (ГЭБ) не происходит спонтанно – как правило, требуется провоцирующий фактор. Много лет, занимаясь лечением шизофрении, мы составили перечень рисков, провоцирующих первый прорыв ГЭБ противомозговыми антителами. Вот ряд таких причин в порядке убывания вреда. Самым гибельным мы считаем курение «спайсов», изобретенных Дж. Хоффманом. Даже однократное курение кальяна с синтетическим аналогом каннабиоида или курительных смесей может оказаться фатальным. То же самое касается марихуаны. Именно прием каннабиоидов для людей, имеющих наследственную предрасположенность к шизофрении, мы полагаем самым опасным провоцирующим моментом. Затем следует черепно-мозговая травма. Затем грипп или респираторная инфекция с высокой температурой. Затем компьютерные игры и интернетзависимость. И, наконец, гормональный сдвиг, сопровождающий период полового созревания. Бывает, что эти факторы накладываются друг на друга. Если сравнить начало шизофрении с боевой гранатой, то провоцирующий фактор можно представить в виде выдернутой чеки. Аутоиммунную атаку на астроциты уподобить запалу. И только потом следует взрыв боевой части в виде бессонницы и разлет осколков в виде псевдогаллюцинаций, бреда, страхов, потери внимания, падения энергии и т.д. Закончится ситуация параноидной или простой формой, аутизмом или гебефренией, шизоаффктивным психозом или кататонией – зависит от наследственного генотипа в виде «мощности заряда и сектора разлета осколков», но принципиально, все типы шизофрении протекают по одному сценарию: нарушение сна (сокращение глубоких фаз медленного сна, изменение латентности REM-сна), компенсаторный «сон» отдельных участков мозга – больше известный как псевдогаллюцинации (сон в бодствовании), бредовые идеи, порожденные псевдогаллюцинациями, падение энергетического потенциала, расстройство внимания и т.д.
Мы утверждаем, что специфические нарушения сна характерны для всех типов шизофрении, включая аутизм. Хочется обратить внимание читателей на поразительное сходство психических расстройств, зарегистрированных у молодых, психически здоровых добровольцев в эксперименте с депривацией сна (ДС) - длительным (больше 200 часов) искусственным лишением сна [32]. В любом психиатрическом стационаре под действием нейролептиков, все пациенты «хорошо спят». Но вопрос о том, приносит ли такой сон желанный отдых, восстановление сил и эмоциональный комфорт – остается открытым. Без специальной аппаратуры отличить нейролептический сон от физиологического практически невозможно. Только полисомнография, которую в стационарах не проводят, позволяет судить о качестве и глубине сна, да и то – частично. Прибор регистрирует электрическую активность мозга, но как на клеточном уровне ведет себя спящий мозг – известно мало.

Рис 1. Полисомнография пациентки Г. на фоне приема нейролептиков (Дельта сон на фоне приема нейролептиков. Эффективность сна – 10%).
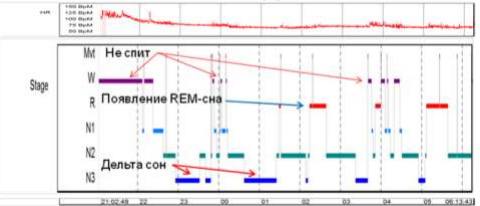
Рис.2. Полисомнография пациентки Г. после курса ингаляций ККРЦ (Дельта сон + REM-сон после 2 месяцев ингаляций ККРЦ. Эффективность сна – 80%).
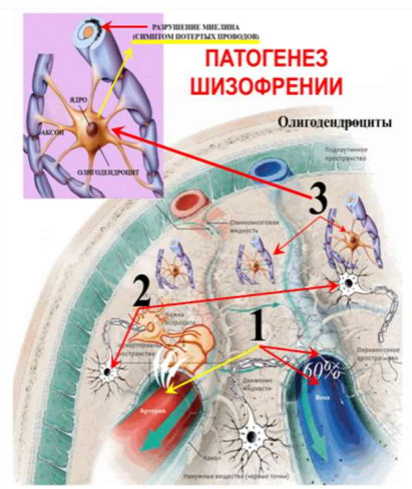
Рис. 3. Патогенез шизофрении (общий план рисунка взят из Журнала «В мире науки». 2016, с изменениями автора). Пояснения: 1 – Аутоиммунная атака антител на «ножки» астроцитов. Нарушение их группового взаимодействия. Торможение глифатической системы. Накопление продуктов метаболизма. 2 – Прекращение энергетической подпитки нейронов. Снижение передачи лактата и митохондрий из астроцитов в нейроны. 3 – Деградация и гибель олигодентроцитов. Демиелинизация аксонов. Симптом «потертых проводов». Рассеивание импульса).
А сон, между тем, в значительной мере организован не только нейронами, но и клетками глии. Во всяком случае, глубокий медленный сон, который еще называют ДЕЛЬТА СОН. В этом смысле полисомнографию можно сравнить с кардиограммой, электрические сигналы которой позволяют частично судить о работе сердца.
Психиатры привычно решают проблему бессонницы назначением психотропных препаратов, часто не подозревая, что лекарства, способного вернуть пациенту с шизофренией физиологический сон пока не создано, а медикаментозный сон – разительно отличается от физиологического. Особенно по количеству дельта сна…
Заглянуть в ткань живого мозга, чтобы понять, как он спит – впервые удалось в 2012 г. Стивену Голдману и Майкену Недергарду при помощи двухфотонной микроскопии. Они следили за распределением специально окрашенного ликвора. Оказалось, что во время дельта сна ликвор распределялся по ткани мозга во много раз быстрее. Так была открыта глимфатическая система большого мозга.
Больше десяти лет назад, в самом начале клинического эксперимента, по лечению шизофрении композиционным раствором цитокинов (ККРЦ), мы были удивлены полученным результатом, но решили понаблюдать, как долго продлится ремиссия. Прошло несколько лет, ремиссия сохранялась… Затем появились другие пациенты, расставшиеся с психозом и нейролептиками с помощью чисто иммунологических методов… Тогда мы сделали первую робкую попытку объяснить патогенез шизофрении, поставив в основу аутоиммунную теорию: «Шизофрения представляет собой органическое поражение мозга, как результат аутоиммунной деструкции нейронов и глии. Клиническая форма и прогредиентность заболевания зависят от локализации, мощности и скорости нарастания деструктивных аутоиммунных процессов».
Через два года, окрыленные хорошими клиническими результатами у следующих пациентов, мы сочли возможным предположить, что «ингалируемая нами терапевтическая смесь цитокинов, сдвигает иммунную реакцию Th2 в сторону Th1 и, таким образом, останавливает разрушительный аутоиммунный процесс».
Одновременно мы предположили, что хоуминг прогенеторных стволовых клеток из обонятельного мозга в очаги дегенерации нейронов – является основным механизмом восстановления ЦНС. Мы полагали, что этот механизм, доказанный Магдаленой Гоц, обеспечивает наблюдаемую нами клиническую ремиссию с редукцией дефекта. Сейчас мы понимаем, что это не совсем так…
Все наши пациенты, прежде чем выйти из психоза в ремиссию, начинали долго и крепко спать. Спали ночью и днем, иногда по 18 часов в сутки… При этом, никаких психотропных препаратов им не давали. Сон был физиологическим. В попытке понять, что происходит – мы стали делать всем пациентам полисомнографию: сразу при поступлении (когда они еще были на больших дозах нейролептиков) и через разные промежутки времени после ингаляций ККРЦ. В результате мы обнаружили, что цитокины являются уникальным инструментом для восстановления естественного, физиологического сна.
В качестве примера приводим две полисомнографии пациентки «Г» 2000 г. рождения. «Г» – инвалид c детства по шизофрении. Госпитализировалась многократно. На нейролептиках регул нет почти год. В очередной раз выписана из стационара на «поддерживающих» дозах галоперидола, кветиопина, арипипразола и амитриптилина одновременно. Со слов мамы, только на этих препаратах может «спать», но меньше 8 часов. Реальная эффективность сна при самом добросовестном употреблении всех психотропных оказалась – 10%. Дельта сна практически нет.
На фоне ингаляций ККРЦ все психотропные препараты постепенно отменены. Через два месяца ингаляций пациентка спит 10-12 часов в сутки, спать ложится рано, сама, и спит порядка 9 часов ночью и пару часов днем. Реальная эффективность сна на повторной полисомнографии – 80%. Самое главное, появился дельта сон! О появлении REM сна следует упомянуть попутно. Появились и стабилизировались регулы, исчезли «голоса», начали сниться сны – вначале кошмарные, а теперь «добрые».
Цифра 1 на Рисунке 3: Двумя красными стрелками, справа показывает увеличенное на 60% глимфатическое пространство в момент «сморщивания» «ножек» астроцитов. После прорыва гематоэнцефалического барьера в ликвор попадают антимозговые антитела. Левая желтая стрелка показывает атаку, попавших в ликвор антимозговых антител на «ножки» астроцитов (три белые стрелки). Атакованные астроциты частично теряют возможность согласованно координировать набухание и сморщивание «ножек» (доменного взаимодействия). Снижается эффективность выведения продуктов метаболизма глимфатической системой. Клинически это выглядит как нарушение сна вплоть до полной бессонницы. На полисомнографии это выглядит как фрагментация, сниженная амплитуда и сокращение времени дельта сна, без возможности его компенсации. Отсутствие возможности компенсации дельта сна разительно отличает сон при всех типах шизофрении от любой другой патологии сна.
Цифра 2 на Рисунке 3 стрелками показывает, что атакованные астроциты теряют способность делится с нейронами собственными митохондриями, но самое главное – падает количество отдаваемого нейронам лактата. Уникальный механизм нейронов, превращающий лактат в 38 молекул АТФ – простаивает без «нефти для бензина». Клинически это выглядит как потеря энергии, астения, аутизм.
Цифра 3 на Рисунке 3 стрелками показывает как поставленные в невыносимые, токсические условия олигодендроциты деградируют и массово гибнут. В результате аксоны нейронов теряют миелиновую изоляцию, что ведет к рассеиванию импульса. Клинически это проявляется целым спектром нарушений мышления, а по прошествии времени приводит к выраженному дефекту.
заболеваний, вызванных "загрязнением" мозга, – считает первооткрыватель глимфатической системы Недергорд. – «Восстановительная функция сна может быть следствием усиленного удаления потенциально нейротоксических отходов, которые накапливаются в бодрствующей ЦНС. Понимание процесса активации глимфатической системы – первый шаг к возможности модулировать эту систему и заставлять её работать эффективнее».
Собственно, гематоэнцефалический барьер и анатомическая структура недавно открытой глимфатической системы – это одно и то же клеточное образование. Разница в названиях: глимфатическая система воспринимается динамичной, поскольку постоянно находится в движении, а гематоэнцефалический барьер кажется статичным, потому что изучался преимущественно на препаратах мозга. Важно понять, что гематоэнцефалический барьер и глимфатическая система представляют собой единый анатомический субстрат, организующий дельта сон.
О существовании гематоэнцефалического барьера знает каждый врач. Эндотелиальные клетки всех кровеносных сосудов мозга настолько плотно прилегают друг к другу, что в периваскулярное пространство из крови не могут попасть молекулы размером больше 1,5 нм (вода, глюкоза). Все это пространство, не менее плотно окружено «ножками» астроцитов. Между эндотелиальной стенкой сосуда и внешней «трубой» из «ножек» астроцитов - есть небольшое, регулируемое астроцитами, пространство, по которому постоянно течет ликвор. Все, что попадает внутрь мозга – проникает туда только через «ножки» астроцитов. Исключением являются анатомические образования: «серый бугор, воронка и задняя доля гипофиза».
Дебют и течение шизофрении практически всегда связаны с бессонницей. Но что такое сон?
На уровне взаимодействия клеток мозга, медленный сон – это, прежде всего, содружественная, скоординированная работа астроцитов, направленная на вымывание из глубин мозга накопившихся метаболитов. Третья и четвертая фазы медленного сна (дельта сон) характеризуются усиленным движением ликвора по периваскулярным пространствам и через ткани мозга. В период бодрствования вымывание метаболитов происходит гораздо медленнее.
В норме, после первой, самой длительной (60минут) фазы дельта сна – наступает РЕМ-фаза, во время которой микроглия движется и «отстригает» неактуальные контакты нейронов (старые синапсы), а когда «отстриженные» контакты накопятся – наступает второй цикл выведения метаболитов. Он уже покороче. И так раз пять за ночь. К утру совсем короткая «уборка» продуктов обмена и, наконец, избавленный от метаболитов мозг, в стадии сна с быстрым движением глаз – восстанавливает все необходимые синапсы. Таким образом, в мозге, как в органе – сон материален и представляет из себя взаимодействие астроцитов, продвигающих ликвор по глимфатической системе, непрерывное движение клеток микроглии, избавляющих нейроны от неиспользуемых контактов, электрическое взаимодействие нейронов, приводящее к новым решениям на основании прежних впечатлений. «Прожектор» сознания в это время сводится к работе сторожевых центров, призванных быстро разбудить мозг в случае опасности (например, пробуждение после апноэ).
Если рассматривать патогенез как цепь событий, приводящих к разбалансировке гармонично протекающих физиологических процессов, то началом шизофрении мы полагаем прорыв гематоэнцефалического барьера, вероятнее всего в области древнего мозга (обонятельный мозг, серый бугор или воронка). Кстати, об этом свидетельствуют обонятельные галлюцинации, нередкие в дебюте шизофрении. Противомозговые антитела попадают в ликвор и атакуют «ножки» астроцитов. Плотно сомкнутые «ножки» астроцитов не только представляют собственно мозговую часть гематоэнцефалического барьера, но они же, совместным доменным набуханием и последующим «сморщиванием» – образуют двигатель глимфатической системы, обеспечивающий своевременное выведение продуктов метаболизма. В результате аутоиммунного поражения астроцитов нарушается согласованная работа их кальциевых каналов, происходит серьезная разбалансировка содружественного «набухания» и «сморщивания» этих «ножек». Клинически это проявляется уменьшением времени и эффективности дельта сна, который обеспечивает эффективность работы глимфатической системы. На полисомнограмме регистрируется снижение амплитуды дельта колебаний, появляется фрагментация дельта сна, сокращается его время.
Только при шизофрении мы регистрируем бессонницу или резкое сокращением дельта сна, с невозможностью его компенсации после депривации. Так не происходит ни при каких нейродегенеративных заболеваниях. Нарушение слаженной работы глимфатической системы приводит к накоплению продуктов метаболизма, самоотравлению мозга, образованию многочисленных пространств Вирхова-Робина и очагов глиоза, хорошо видных на ЯМР. Дисгармония доменного взаимодействия астроцитов, как основных организаторов дельта сна, приводит к сепаратным попыткам отдельных участков мозга (преимущественно височных областей) спать в период общего бодрствования, на фоне полного сознания отдельными участками – (пирамидками). Такое же явление наблюдается при длительной депривации сна у совершенно здоровых людей. У них появляются «сновидения» на фоне бодрствования в виде псевдогаллюцинаций.
Такие «псевдосновидения» могут регистрироваться энцефалограммой как сонные веретена и даже К-комплексы прямо днем, во время бодрствования (на круглосуточной полисомнографии высокого разрешения). Кроме того, пораженные астроциты перестают делится митохондриями и лактатом с нейронами. Клинически это проявляется общим упадком энергии, астенией, «обломовской ленью».
Накопление продуктов метаболизма в результате сбоя в работе глимфатической системы, ведет к массовой гибели олигодендроцитов. Электронная микроскопия пирамидного слоя гиппокампа у больных шизофренией выявила патологические включения между слоями миелиновой оболочки большинства миелинизированных аксонов.
Возникает так называемый – «симптом потертых проводов». Это приводит к «расползанию, деканализации» генерируемых импульсов, что клинически выражается разнообразными расстройствами мышления – философическая интоксикация, рассеянность, трудность концентрации, в значительной степени нарушается внимание.
Чем меньше дельта сна – тем ярче клиника. Органическое поражение мозга нарастает по мере увеличения количества приступов, а при непрерывном течении, вместе с длительностью периода отсутствия дельта сна. Шизофрения представлена прежде всего морфологическим субстратом нейровоспаления и нейродегенерацией глии. Нейроны страдают во вторую очередь.
Мозг как орган материален, состоит из клеток, которые обмениваются митохондриями, микроглия перемещаются между нейронами, отстригает «лишние» синапсы, набухают и опадают «ножки» астроцитов, перетекает по периваскулярным пространствам ликвор и т.д. Электрические потенциалы, полисомнографии говорят о его работе только косвенно. Сомнолог Сейпер в 2005 году опубликовал в журнале Nature концепцию «переключения» сна и бодрствования. Чем активней бодрствование, тем с большей силой «центр засыпания» испытывает потребность щелкнуть переключателем на сон. Так называемый «флип флоп». Чем дольше сон, тем меньше потребность в сне. Так что в какой-то момент система бодрствования берёт верх. Мы просыпаемся и чувствуем, что выспались.
Такой подход не выдерживает критики. Мозг в норме стремится спать постоянно, во всяком случае до тех пор, пока внутренний или внешний стимул не возбудят центры бодрствования. Это ядра ретикулярной формации, каждое из которых возбуждается собственным медиатором, а их как минимум – семь. Возбуждаются они только все вместе. Выход из строя даже одного ядра – несовместим с сознанием. Бодрствование наступает, когда появляется внутренний или внешний стимул; например жажда или изменение окружающей температуры. И никакого резкого перехода от сна к бодрствованию не существует. Сон никогда не возникает мгновенно, а прежде чем окончательно проснуться мозг проходит через ряд просоночных состояний. Псевдогаллюцинации при шизофрении и при длительной депривации сна у здоровых – это и есть «сны наяву». Концепция Спейсера не объясняет что такое дремота или кратковременное засыпание (за рулем). Это сон или резкое падение внимания? А сноговорение, снохождение и т.д.?
Полное лишение возможности спать у здоровых людей, может привести к спутанности сознания, галлюцинациям, эпилептическим припадкам и даже смерти. Доказано, что длительная депривация сна приводит к гибели нейронов. Лабораторные животные погибают, если их лишать сна в течение нескольких дней, а люди не намного устойчивее животных [18]. Пациенты с диагнозом шизофрения, не имеют полноценного сна годами. Продукты метаболизма накапливаются постоянно, с весьма ограниченной возможностью когда-либо покинуть мозг. Вот Вам и дефект, на который нейролептики повлиять не могут.
На основе всего вышеизложенного мы предположили, что нарушения сна при шизофрении могут быть не побочным эффектом, а ведущим механизмом болезни. В основе патогенеза шизофрении лежит специфическая патология сна, которую современные психотропные средства устранить не в состоянии. Специфика состоит в том, что после лишения сна у всех здоровых добровольцев и при всех иных видах патологии, включая депрессию, в первую очередь компенсаторно удлиняется фаза глубокого медленного сна. Мозг старается как можно быстрее избавиться от продуктов метаболизма. При шизофрении этого не происходит. Количество и качество дельта сна после депривации остается прежним. Никакой компенсации нет. Другими словами, общаясь с «лично больным» Вы имеете дело с человеком, который не способен полноценно спать годами. Его мозг начинает спать участками (пирамидками), сонные веретена и Ккомплексы появляются прямо на фоне бодрствования, хотя какое это «бодрствование», каждый может почувствовать на себе, если не будет спать хотя бы неделю, а лично больные в таком состоянии находятся годами. Компенсаторный сон участков мозга в период бодрствования – это и есть псевдогаллюцинации, (голоса, полупрозрачные видения) характерные исключительно для шизофрении.
Заключение.
Основной принцип, вокруг которого строится патогенез шизофрении, действительно лежит на поверхности – это специфическая патология сна. Недостаток дельта сна снижает возможности глимфатической системы выводить продукты метаболизма. Это приводит к хроническому самоотравлению мозга, гибели отдельных видов микроглии, потере энергетической подпитки нейронов, гибели олигодендроцитов, диемиелинизации аксонов. Шизофрения прежде всего болезнь глии! Нейроны страдают вторично и долгое время сохраняют возможность практически полного восстановления.
Эксперимент продолжается. Сейчас мы имеем пациентов с диагнозом F20, у которых после применения цитокинов, произошла редукция дефектной симптоматики и много лет наблюдается стойкая клиническая ремиссия. Они живут самостоятельно, работают, учатся, рожают детей, и в поддерживающем лечении не нуждаются. При этом мы понимаем, что возможность возникновения нового обострения сохраняется постоянно. От генетической предрасположенности никуда не уйдешь. Поэтому мы не рекомендуем нашим пациентам чрезмерно напрягать «ограниченные возможности» своего мозга. Стараться меньше смотреть в компьютер, не работать в ночные смены, больше времени проводить на природе, избегать травм, ни в коем случае не употреблять канабиоиды.
Литература:
1. Бехтерев В.М. Будущее психиатрии. СПб.: Наука, 1997. С. 308.
2. Бредбери М. Концепция гематоэнцефалического барьера. М.: М, 1983. С. 90-130.
3. Ветлугина Т.П. Иммунная система при шизофрении. Томск: МГП «РАСКО», 2000. 112 с.
4. Ветлугина Т.П. Психонейроиммуномодуляция при шизофрении. Возможности прогноза течения и терапии на основе иммунологических подходов. В кн: Биологические маркеры шизофрении: поиск и клиническое применение / Под ред. Н. А. Бохана, С.А. Ивановой. Новосибирск: Изд-во СО РАН, 2017. С. 23-33.
5. Воронов А. И. Шизофрения (болезнь Блейлера) между двумя юбилеями // Академический журнал Западной Сибири. 2010. № 2. С. 10-13.
6. Воронов А.И., Дресвянников В.Л., Пухкало К.В. Шизофрения. Новый способ лечения // Тюменский медицинский журнал. 2012. № 1. С. 9-15.
7. Коломеец Н.С. Межклеточные взаимодействия в мозге человека при шизофрении (ультраструктурно – морфометрическое исследование): Дисс.. докт. биол. наук. Москва, 2010. 110 с.
8. Коляскина Г.И. Иммунология в психиатрии. В кн.: Руководство по психиатрии. Под ред. А.С. Тиганова. М, 1999. Т. 1. С. 190 211.
9. Кутько И.И., Фролов В.М., Рачкаускас Г.С. Детоксикация и коррекция метаболических расстройств в лечении больных шизофренией // Психиатрия. 2006. № 1. С. 26-30.
10. Осокина О. И. Нейроанатомические и гистологические предпосылки шизофрении (обзор проблемы) // Український вісник психоневрології. 2016. Т. 24, № 4. С. 89.
11. Самохина Е. «Прожигатель» энергии // Наука и жизнь. 2017. № 4. С. 5-7.
12. Семке В.Я., Ветлугина Т.П., Невидимова Т.А., Иванова С.А., Бохан Н.А. Клиническая психонейроиммунология. Томск: Издво «РАСКО», 2003. 300 с.
13. Снесарев П.Е. Теоретические основы патологической анатомии психических болезней. М.: МЕДГИЗ, 1950. 370 с.
14. Руководство по психиатрии / Под ред. Тиганова А.С. 1999. Ч. 2. 265 с.
15. Чуркин А.А., Матюшов А.Н. Практическое руководство по применению МКБ-10 в психиатрии и наркологии. М, 2010. С. 48-58.
16. Araque A., Parpura V. et al. Tripartite synapses: glia, the unacknowledged partner // Trends Neurosci. 1999. № 22. P. 208–215.
17. Bacaj T., Tevlin M., Lu Y., Shaham, S. Glia are essential for sensory organ function in C. elegans // Science. 2008. Vol. 322. P. 744–747.
18. Bryan F. Sleep deprivation – Better Health Channel Sleep Deprivation Proc // Natl Acad Sci USA. 2009. Sep 1. Vol. 106. P. 35.
19. Carlsson A., Rosengren E., Bertler Å. and Nilsson J. Internat. Symp. On Psychotropic Drugs // Nature. 1957. May. Р. 9–11.
20. Fellin T. et al. Endogenous nonneuronal modulators of synaptic transmission control cortical slow oscillations in vivo // Proc. Natl Acad. Sci. USA. 2009. Vol. 106. P. 15037-15042.
21. Ferrarelli F., Tononi G. Reduced sleep spindle activity point to a TRN-MD thalamus-PFC circuit dysfunction in schizophrenia // Schizophr Res. 2017. T. 180. C. 36-43.
22. Frank O., Giehl M., Zheng C. et al. Human endogenous retrovirus expression profiles in samples from brains of patients with schizophrenia and bipolar disorders // J. Virology. 2005. Vol. 79 (17). P. 10890-10901.
23. Gotz Magdalena Progress in Exploring New Avenues for Brain Repair: Astroglia Reprogrammed to Generate Synapse-Forming Neurons // https://www.helmholtzmuenchen.de/isf/service/news/news/article/19 968/index.html
24. Hayakawa K, Esposito E. et al. Transfer of mitochondria from astrocytes to neurons after stroke // Nature. 2016. July. P. 28.
25. Karlsson H., Bachmann S.J., Schroder J. et al. Retroviral RNA identified in the cerebrospinal fluids and brains of individuals with schizophrenia // Proc Nat Acad Sci USA. 2001. Vol. 98. P. 4634-4639.
26. Muller N., Riedel M., Ackenheil M., Schwarz M.J. Cellular and humoral immune system in schizophrenia: a conceptual reevaluation // World J Biol Psychiat. 2000. Vol. 1. P. 173-179.
27. Nikkila H.V., Muller K., Ahokas A. et al. Accumulation of macrophages in the CSF of schizophrenic patients during acute psychotic episodes // Am J Psychiat. 1999. Vol. 156 (11). P. 1725-1729.
28. Nikkila H.V., Muller K., Ahokas A. et al. Increased frequency of activated lymphocytes in the cerebrospinal fluid of patients with acute schizophrenia // Schizophr Res. 2001. Vol. 49 (1-2). P. 99-105.
29. Olson J.K., Miller S.D. Microglia initiate central nervous system innate and adaptive immune responses through multiple TLRs // J Immunol. 2004. Vol. 173. P. 3916-3924.
30. Perea G., Araque A. Astrocytes potentiate transmitter release at single hippocampal synapses // Science. 2007. Vol. 317. P. 1083–1086.
31. Pia Rivetti di Val Cervo et al. Induction of functional dopamine neurons from human astrocytes in vitro and mouse astrocytes in a Parkinson’s disease model // Nature Biotechnology. 2017. doi:10.1038/nbt.3835
32. Purves D., Augustine G.J., Fitzpatrick D., et al.. Why Do Humans and Many Other Animals Sleep? // Neuroscience. 2001. № 2.
33. Rosenhan D.L. On being sane in insane places // Science. (New York, N.Y.). January. 1973. № 179 (70). P. 250-258.
34. Steven A. Goldman, Nedergaard Maiken The Brain's WasteDisposal System May Be Enlisted to Treat Alzheimer's and Other Brain Illnesses // Sientific American. 2016. March. 1.
35. Strous R.D., Shoenfeld Y. Schizophrenia, autoimmunity and immune system dysregulation: a comprehensive model updated and revisited // J Autoimmun. 2006. Vol. 27 (2). P. 71-80.
36. Town T., Nikolic V., Tan J. The microglial “activation” continuum: from innate to adaptive responses // J Neuroinflammation. 2005. Vol. 2. P. 24.
37. Welberg L. Neuronglia interactions: Glia make waves // Nature Reviews Neuroscience. 2009. September. 9.